Submissions





GlyTouCan
Glycan Structure Repository
GlyComb
Glycoconjugate Repository
GlycoPOST
Glycomics MS raw data Repository
UniCarb-DR
Glycomics MS Repository for glycan annotations from GlycoWorkbench
LM-GlycoRepo
Repository for lectin-assisted multimodality dataAll Resources
Genes / Proteins / Lipids Glycans / Glycoconjugates Glycomes Pathways / Interactions / Diseases / OrganismsTools
Guidelines
MIRAGE G57321FI
G57321FI
Summary
- GlyTouCan ID
-
G57321FI
- IUPAC Condensed
- GalNAc(a1-
- Motifs
- Tn antigen
Organisms
| Organisms | Evidence |
|---|---|
| Ovis aries (sheep) | |
| Cricetulus griseus (Chinese hamster) | |
| Mus musculus (house mouse) | |
| Bos taurus (domestic cattle) | |
| Escherichia coli |
Taxonomic Hierarchy
root
cellular organisms [inference]
Eukaryota (eucaryotes) [inference]
Opisthokonta [inference]
Metazoa (metazoans) [inference]
GlycoGene Database (GGDB)
| Gene Symbol | Donor | Acceptor | Reducing terminal(Acceptor) | Product | Reducing terminal(Product) | Reference |
|---|---|---|---|---|---|---|
| C1GALT1 | (not applicable) |
 G57321FI
G57321FI
|
Ser/Thr |
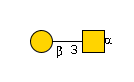 G00031MO
G00031MO
|
Ser/Thr | |
| ST6GALNAC1 | CMP-Neu5Ac |
G57321FI
|
Ser/Thr |
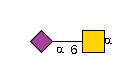 G36123IU
G36123IU
|
Ser/Thr | |
| ST6GALNAC1 | CMP-Neu5Ac |
G57321FI
|
O-glycan Synthesis |
G36123IU
|
O-glycan Synthesis | |
| B3GNT6 | UDP-GlcNAc |
G57321FI
|
[alpha]-pNP |
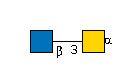 G00035MO
G00035MO
|
[alpha]-pNP |
| Gene Symbol | Donor | Acceptor | Reducing terminal(Acceptor) | Product | Reducing terminal(Product) | Reference |
|---|---|---|---|---|---|---|
| C1GALT1 | (not applicable) |
G57321FI
|
Ser/Thr |
G00031MO
|
Ser/Thr | |
| ST6GALNAC1 | CMP-Neu5Ac |
G57321FI
|
Ser/Thr |
G36123IU
|
Ser/Thr | |
| ST6GALNAC4 | CMP-Neu5Ac |
G57321FI
|
benzyl |
G36123IU
|
benzyl | |
| ST6GALNAC1 | CMP-Neu5Ac |
G57321FI
|
O-glycan Synthesis |
G36123IU
|
O-glycan Synthesis | |
| ST6GALNAC1 | CMP-Neu5Ac |
G57321FI
|
[Ala-Thr(*)-Ala]2-7 |
G36123IU
|
[Ala-Thr(*)-Ala]2-7 |
| Gene Symbol | Donor | Acceptor | Reducing terminal(Acceptor) | Reference |
|---|---|---|---|---|
| ST6GAL1 | CMP-Neu5Ac |
G57321FI
|
para-nitrophenol | |
| ST8SIA6 | CMP-Neu5Ac |
G57321FI
|
||
| ST6GALNAC2 | CMP-Neu5Ac |
G57321FI
|
para-nitrophenol | |
| GAL3ST2 | PAPS |
G57321FI
|
[alpha]-benyzl | |
| B4GALT2 | UDP-Gal |
G57321FI
|
Benzyl-[alpha] |
KEGG BRITE Database
 G00041MO
G00041MO
Core Protein
GlycoEpitope
- Epitope ID
- EP0021
Pathway
| Pathway Name | Organism |
|---|---|
| E.coli O5ab | Escherichia coli |
| E.coli O5ac | Escherichia coli |
| E.coli O61 | Escherichia coli |
| E.coli O65 | Escherichia coli |
| E.coli O6H10K54 | Escherichia coli |
| E.coli O6H16K15 | Escherichia coli |
| E.coli O70 | Escherichia coli |
| E.coli O71 | Escherichia coli |
| E.coli O73ab | Escherichia coli |
| E.coli O86 | Escherichia coli |
Sequence Descriptors
- GlycoCT
-
RES 1b:a-dgal-HEX-1:5 2s:n-acetyl LIN 1:1d(2+1)2n
- WURCS
- WURCS=2.0/1,1,0/[a2112h-1a_1-5_2*NCC/3=O]/1/
Literature
| PubMed ID | Title | First Author | Publication Date | Source |
|---|---|---|---|---|
| 21799473 | Substrate Promiscuity of N-Acetylhexosamine 1-Kinases | Li Y | 2011 Jul 28 |
|
| 21999536 | Structural diversity of the core oligosaccharide domain of Pseudomonas aeruginosa lipopolysaccharide | Kocincova D | 2011 Jul |
|
| 21440051 | Bacterial adherence in otitis media: Determination of N-acetylgalactosamine (GalNAc) residues in the submucosal glands and surface epithelium of the normal and diseased Eustachian tube | Kirkeby S | 2011 Jul |
|
| 20473945 | Tumour‐associated glycan modifications of antigen enhance MGL2 dependent uptake and MHC class I restricted CD8 T cell responses | Singh SK | 2011 Jan 28 |
|
| 20934911 | A simple fibril and lectin model for cyst walls of Entamoeba and perhaps Giardia | Samuelson J | 2011 Jan |
|
| 21307569 | Bacillus thuringiensisCry Toxins Bound Specifically to Various ProteinsviaDomain III, Which Had a Galactose-Binding Domain-Like Fold | KITAMI M | 2011 Feb 23 |
|
| 21358053 | Modification of the sugar specificity of a plant lectin: structural studies on a point mutant ofErythrina corallodendronlectin | Thamotharan S | 2011 Feb 12 |
|
| 21138417 | Chondroitin 4-O-sulfotransferase-1 regulates the chain length of chondroitin sulfate in co-operation with chondroitin N-acetylgalactosaminyltransferase-2 | Izumikawa T | 2011 Feb 11 |
|
| 21303815 | Structure of a lectin with antitumoral properties in king bolete (Boletus edulis) mushrooms | Bovi M | 2011 Feb 08 |
|
| 21300897 | Metabolic cross-talk allows labeling of O-linked beta-N-acetylglucosamine-modified proteins via the N-acetylgalactosamine salvage pathway | Boyce M | 2011 Feb 07 |
|
PubAnnotation

